Схема синтеза соляной кислоты в желудке

Оглавление темы “Глотание. Пищеварение в желудке.”:
1. Глотание. Акт глотания. Фазы глотания. Пищевой комок.
2. Пищеварение в желудке. Функции желудка.
3. Секреторная функция желудка. Желудочный сок. Состав желудочного сока.
4. Соляная кислота. Механизм секреции соляной кислоты. Образование соляной кислоты в желудке.
5. Роль соляной кислоты в пищеварении. Функции соляной кислоты. Ферменты желудочного сока и их роль в пищеварении.
6. Желудочная слизь и ее значение. Слизь желудка. Функции желудочной слизи.
7. Регуляция секреции желудочного сока. Принципы секреции желудочного сока.
8. Фазы желудочной секреции. Нейрогуморальная фаза. Кишечная фаза.
9. Желудочная секреция при переваривании различных пищевых веществ. Секреция на белки. Секреция на углеводы. Секреция на молоко.
10. Сократительная деятельность мускулатуры желудка. Сокращение желудка. Работа желудка.
Соляная кислота. Механизм секреции соляной кислоты. Образование соляной кислоты в желудке.
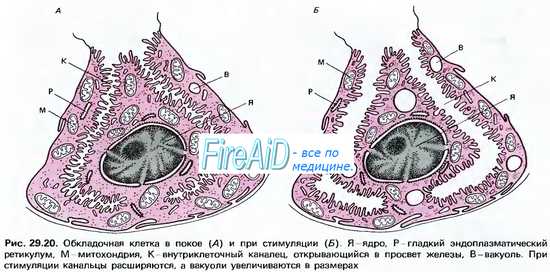
Хлористоводородная кислота вырабатывается париетальными (обкладочными) клетками желез желудка. Эти клетки характеризуются богатством митохондрий, расположенных вдоль внутриклеточных канальцев. Площадь мембраны канальцев и апикальной поверхности клеток во время стимуляции на высоте секреции резко возрастает за счет встроенных в мембрану тубовезикул (трубочек-пузырьков), что сопровождается значительным увеличением клеточных канальцев, проникающих вплоть до базальной мембраны. Это значительно увеличивает возможности синтеза гландулоцитом соляной кислоты. Вдоль канальцев располагается множество митохондрий, площадь внутренней мембраны которых возрастает в процессе биосинтеза НСl. Соответственно увеличивается площадь контакта канальцев и апикальной мембраны клетки. Таким образом, увеличение секреторной активности париетальных клеток обусловлено увеличением площади секреторной мембраны.
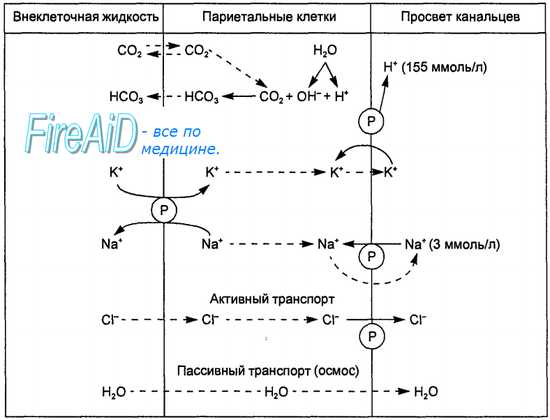
Рис. 11.11. Образование соляной кислоты желудочного сока. Пояснения в тексте.
Символ ® означает активность ферментных транспортных систем мембраны кислотопродуцирующих клеток. Стрелками показано направление движения ионов и воды.
Секреция НСl является ярко выраженным цАМФ-зависимым процессом, активация которого протекает на фоне усиления гликогенолитиче-ской и гликолитической активности, что сопровождается продукцией пирувата. Окислительное декарбоксилирование пирувата до ацетил-КоА. С02 осуществляется пируватдегидрогеназным комплексом и сопровождается накоплением в цитоплазме НАД • Н2. Последний используется для генерирования Н+ в процессе секреции НС1. Расщепление триглицеридов в слизистой оболочке желудка под влиянием триглицеридлипазы и последующая утилизация жирных кислот создает в 3—4 раза больший приток восстановительных эквивалентов в митохондриальную цепь переноса электронов. Как аэробный гликолиз, так и окисление жирных кислот запускаются посредством цАМФ-зависимого фосфорилирования соответствующих ферментов, обеспечивающих генерирование ацетил-КоА в цикле Кребса и восстановительных эквивалентов для электронпереносящей цепи митохондрий. Са2+ является необходимым элементом секреторной системы НС1.
Процесс цАМФ-зависимого фосфорилирования обеспечивает активацию желудочной карбоангидразы, которая является регулятором кислотно-основного равновесия в кислотопродуцирующих клетках. Работа этих клеток сопровождается длительной и массовой потерей ионов Н+, что приводит к накоплению в клетке ОН”, способных оказать повреждающее действие на клеточные структуры. Нейтрализация гидроксильных ионов и является главной функцией карбоангидразы. Образующиеся бикарбонатные ионы посредством электронейтрального механизма выводятся в кровь, а ионы СГ входят в клетку.
Кислотопродуцирующие клетки на наружных мембранах имеют две мембранные ферментные системы, участвующие в механизмах продукции Н+ и секреции НС1. Ими являются Na+-K+-ATФaзa и Н+-К+-АТФаза. Na+-К+-АТФаза, расположенная в базолатеральных мембранах клеток, переносит К+ из крови в обмен на Na+, а Н+-К+-АТФаза, локализованная в секреторной мембране, транспортирует калий из первичного секрета в обмен на выводимые в желудочный сок ионы Н+. Процесс образования соляной кислоты кислотопродуцирующими клетками схематически представлен на рис. 11.11.
В период секреции митохондрии всей массой охватывают в виде муфты секреторные канальцы, и их мембраны сливаются, образуя митохондриально-секреторный комплекс, где ионы Н+ непосредственно акцептируются Н+-К+-АТФазой секреторной мембраны и транспортируются из клетки.
Таким образом, кислотообразующая функция обкладочных клеток осуществляется благодаря процессу фосфорилирования — дефосфорилирования, наличию митохондриальной окислительной цепи, транспортирующей ионы Н+ из матриксного пространства, а также активности Н+-К+-АТФазы секреторной мембраны, перекачивающей протоны из клетки за счет энергии АТФ.
Вода поступает в канальцы клетки путем осмоса. Конечный секрет, поступающий в канальцы, содержит НСl в концентрации 155 ммоль/л, хлористый калий в концентрации 15 ммоль/л и очень малое количество хлористого натрия.
– Также рекомендуем “Роль соляной кислоты в пищеварении. Функции соляной кислоты. Ферменты желудочного сока и их роль в пищеварении.”
Источник
Расщепление белков до аминокислот начинается в желудке, продолжается в двенадцатиперстной кишке и заканчивается в тонком кишечнике. В некоторых случаях распад белков и превращения аминокислот могут происходить также в толстом кишечнике под влиянием микрофлоры.
Протеолитические ферменты подразделяют по особенности их действия на экзопептидазы, отщепляющие концевые аминокислоты, и эндопептидазы, действующие на внутренние пептидные связи.
В желудке пища подвергается воздействию желудочного сока, включающего соляную кислоту и ферменты. К ферментам желудка относятся две группы протеаз с разным оптимумом рН, которые упрощенно называют пепсин и гастриксин. У грудных детей основным ферментом является реннин.
Регуляция желудочного пищеварения
Регуляция осуществляется нервными (условные и безусловные рефлексы) и гуморальными механизмами. К гуморальным регуляторам желудочной секреции относятся гастрин и гистамин.
Гастрин секретируется специфичными G-клетками пилорического отдела:
- в ответ на раздражение механорецепторов,
- в ответ на раздражение хеморецепторов (продукты первичного гидролиза белков),
- под влиянием n.vagus.
Далее гастрин через системный кровоток достигает и стимулирует главные, обкладочные и добавочные клетки, что вызывает секрецию желудочного сока, в большей мере соляной кислоты. Также он обеспечивает секрецию гистамина, влияя на ECL-клетки (enterochromaffin-like cells, англ. энтерохромаффиноподобные клетки).
Гистамин, образующийся в энтерохромаффиноподобных клетках слизистой оболочки желудка (фундальные железы), выходит в кровоток, взаимодействует с Н2-рецепторами на обкладочных клетках и увеличивает в них синтез и секрецию соляной кислоты.
Закисление желудочного содержимого (pH 1,0) по механизму обратной отрицательной связи подавляет активность G-клеток, снижает секрецию гастрина и желудочного сока.
Соляная кислота
Одним из важнейших компонентов желудочного сока является соляная кислота. В образовании соляной кислоты принимают участие париетальные (обкладочные) клетки желудка, секретирующие ионы Н+. Источником ионов Н+ является угольная кислота, образуемая ферментом карбоангидразой. При ее диссоциациии , кроме ионов водорода, образуются карбонат-ионы НСО3–. Они по градиенту концентрации движутся в кровь в обмен на ионы Сl–. В полость желудка ионы Н+ попадают энергозависимым антипортом с ионами К+ (Н+,К+-АТФаза), хлорид-ионы перекачиваются в просвет желудка также с затратой энергии.
Н+,К+-АТФаза (протонная помпа) является мишенью действия лекарственных препаратов “ингибиторов протонной помпы” – омепразол, пантопразол и др., используемых для лечения заболеваний желудочно-кишечного тракта, связанных с повышенной кислотностью (гастриты, язвы желудка и 12-перстной кишки, дуоденит).
При нарушении нормальной секреции HCl возникают гипоацидный или гиперацидный гастрит, отличающиеся друг от друга по клиническим проявлениям, последствиям и требуемой схеме лечения.
Синтез соляной кислоты
Функции соляной кислоты
- Денатурация белков пищи.
- Бактерицидное действие.
- Высвобождение железа из комплекса с белками, что необходимо для его всасывания. Аналогично высвобождаются и другие металлы.
- Высвобождение различных органических молекул, прочно связанных с белковой частью (гем, коферменты – тиаминдифосфат, ФАД, ФМН, пиридоксальфосфат, кобаламин, биотин), что позволяет витаминам впоследствии всасываться.
- Превращение неактивного пепсиногена в активный пепсин.
- Снижение рН желудочного содержимого до 1,5-2,5 и создание оптимума рН для работы пепсина.
- После перехода в 12-перстную кишку – стимуляция секреции кишечных гормонов и, следовательно, выделения панкреатического сока и желчи.
Кислая реакция желудочного сока обусловлена, главным образом, присутствием HCl, гораздо в меньшей степени иона H2PO4-, при патологиях (гипо- и анацидное состояние, онкология) свой вклад может вносить молочная кислота.
Совокупность всех веществ желудочного сока, способных быть донорами протонов, составляет общую кислотность. Соляную кислоту, находящуюся в комплексе с белками, мукополисахаридами слизистой оболочки и продуктами переваривания, называют связанной соляной кислотой, оставшуюся часть – свободной соляной кислотой. Содержание свободной HCl подвержено изменениям, в то же время количество связанной HCl относительно постоянно.
Влияние гастрина и гистамина на обкладочные клетки сводится к усилению работы Н+,К+-АТФазы. Действие гастрина заключается в активации кальций-фосфолипидного механизма передачи сигнала, гистамин действует по аденилатциклазному механизму.
Изменение кислотности в желудке
Гипоацидное состояние развивается при снижении активности и/или количества обкладочных клеток, синтезирующих HCl. В результате могут развиваться самые разнообразные последствия, прямо или косвенно связанные с невыполнением соляной кислотой ее функций:
- снижение переваривания белков как в желудке, так и в кишечнике,
- активация процессов брожения в желудке, запах изо рта,
- активация процесса гниения белков в толстой кишке, бурление в кишечнике и метеоризм,
- проникновение недопереваренных продуктов в кровь и, как следствие, аллергические реакции,
- уменьшение высвобождения от белков и возникновение дефицита минеральных веществ (железо, медь, магний, цинк, йод и др),
- снижение высвобождения от белков и всасывания ряда водорастворимых витаминов – развитие гиповитаминозов (B1, B2, B6, B12, H),
- снижение синтеза обкладочными клетками внутреннего фактора Касла и снижение всасывания витамина B12,
- снижение секреции кишечных гормонов и, как следствие, уменьшение выделения желчи и панкреатического сока,
- нарушение переваривания и всасывания липидов и, как следствие, развитие гиповитаминозов по жирорастворимым витаминам.
Гиперацидное состояние развивается при повышенной активности обкладочных клеток. Может приводить к клиническим проявлениям в виде воспаления стенки желудка, эрозии и язвенной болезни желудка и двенадцатипеперстной кишки.
Пепсин
Пепсин является эндопептидазой, то есть он расщепляет внутренние пептидные связи в молекулах белков и пептидов. Синтезируется в главных клетках желудка в виде неактивного профермента пепсиногена, в котором активный центр “прикрыт” N-концевым фрагментом. При наличии соляной кислоты конформация пепсиногена изменяется таким образом, что “раскрывается” активный центр фермента, который отщепляет остаточный пептид (N-концевой фрагмент), т.е. происходит аутокатализ. В результате образуется активный пепсин, активирующий и другие молекулы пепсиногена.
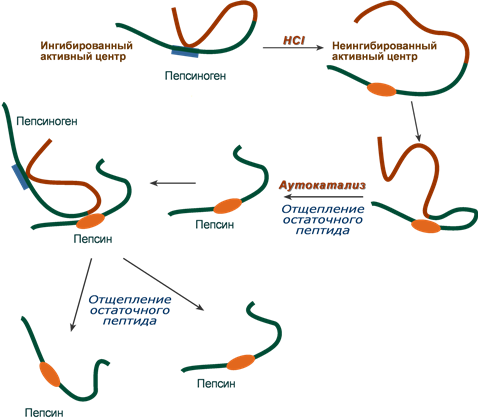
Превращение пепсиногена в пепсин
Пепсин обладает невысокой специфичностью, в основном он гидролизует пептидные связи, образованные аминогруппами ароматических аминокислот (тирозина, фенилаланина, триптофана), меньше и медленнее – аминогруппами и карбоксигруппами лейцина, глутаминовой кислоты и т.д. Оптимум рН для работы пепсина 1,5-2,0.
Связи, расщепляемые пепсином
Гастриксин
Гастриксин по своим функциям близок к пепсину, его количество в желудочном соке составляет 20-50% от количества пепсина. Синтезируется главными клетками желудка в виде прогастриксина (профермент) и активируется соляной кислотой. Оптимум рН гастриксина соответствует 3,2-3,5 и значение этот фермент имеет при питании молочно-растительной пищей, слабее стимулирующей выделение соляной кислоты и одновременно нейтрализующей ее в просвете желудка. Гастриксин является эндопептидазой и гидролизует связи, образованные карбоксильными группами дикарбоновых аминокислот.
В течение суток синтезируется около 2 г пепсина. Объем работы пепсина составляет примерно 10% от всех пептидных связей белков, попадающих в желудок.
Наличие в желудке двух протеаз, действующих при различных pH, позволяет организму пепсином переваривать белки мясной пищи, стимулирующей секрецию HCL, а гастриксином – белки растительно-молочной пищи.
Источник
Молекулярные механизмы регуляции секреции соляной кислоты слизистой оболочки желудка
О.Д. Лопина, А.А. Котлобай, А.М. Рубцов
Московский государственный университет им. М.В. Ломоносова
Секреция соляной кислоты слизистой оболочкой желудка обеспечивается париетальными (обкладочными) клетками, находящимися в эпителиальном слое желудочных желез фундального отдела. Характерной особенностью этих клеток является присутствие специальных структур, так называемых внутриклеточных канальцев, образованных выпячиваниями апикальной мембраны. Поверхность канальцев, как и поверхность апикальной мембраны, покрыта многочисленными микроворсинками. Благодаря наличию внутриклеточных канальцев и микроворсинок значительно увеличивается поверхность, через которую осуществляется секреция соляной кислоты.
Активация секреции соляной кислоты происходит под действием секретогенов: гистамина, гастрина и ацетилхолина. Она сопровождается существенными морфологическими изменениями париетальных клеток: наблюдается значительное увеличение размеров внутриклеточных канальцев и длины микроворсинок, что приводит к увеличению поверхности мембраны, обеспечивающей секрецию. Кроме того, в активированных париетальных клетках внутриклеточные канальцы открываются в люминальное пространство, что обеспечивает доступ выделяющейся соляной кислоты в просвет желудка.
.")
Рис. 1. Транспортные системы париетальной клетки, обеспечивающие секрецию соляной кислоты (схема).
В проникновении соляной кислоты через апикальную мембрану участвует многокомпонентная транспортная система (рис. 1). Основным элементом этой системы является протонный насос, обеспечивающий АТФ-зависимый обмен внутриклеточных Н+ на внеклеточные К+ [7]. Оба иона переносятся против электрохимического градиента. Из клетки К+ выходят по градиенту, по-видимому, через специальный канал, причем выход этого катиона сопровождается выходом из клетки Cl-. Таким образом, суммарным результатом работы этой транспортной системы являются секреция соляной кислоты в люминальное пространство и циклическое перемещение ионов калия из клетки наружу и в обратном направлении. Cl- входят в клетку через базолатеральную мембрану. В транспорте этого аниона принимает участие НСО3-, /Cl- -анионный обменник. Необходимые для такого обмена НСО3– образуются в клетке в результате работы специального фермента карбоангидразы, обеспечивающего синтез Н2СО3 из углекислого газа, который появляется в клетке в результате метаболических процессов, и воды. Н+, образующийся при диссоциации Н2СО3, секретируется протонным насосом в люминальное пространство. Карбоангидраза локализована в клетке в непосредственной близости от системы внутриклеточных канальцев. При интенсивной работе насоса, когда начинает ощущаться нехватка Н+ внутри клетки, в процесс включаются также встроенные в базолатеральную мембрану катионобменники (К+/Н+ или Na+/Н+), обменивающие внеклеточные Н+ на внутриклеточные К+ или Na+. Таким образом, присутствие дополнительных переносчиков, находящихся на базолатеральной мембране, обеспечивает трансэпителиальный транспорт Cl- и частично Н+.
Роль протонного насоса в системе, обеспечивающей секрецию соляной кислоты, выполняет Н+, К+-АТФаза – фермент, относящийся к семейству АТФаз Р-типа [7]. Ближайшим “родственником” этого фермента является Na+, К+-АТФаза, которая вместе с Н+, К+-АТФазой образует отдельное подсемейство. Кроме эпителиальных клеток желудка, Н+, К+-АТФаза (по-видимому, ее изозим) встречается также в эпителиальных клетках почечных канальцев и в эпителии некоторых отделов кишечника.
Н+, К+-АТФаза локализована в апикальной мембране, тогда как Na+, К+-АТФаза сосредоточена исключительно в базолатеральной мембране. Как и Na+, К+-АТФаза, Н+, К+-АТФаза состоит из субъединиц двух типов: a-субъединицы – полипептида с молекулярной массой около 100 кДа (1033 аминокислотных остатка), выполняющего каталитическую функцию, и b-субъединицы – гликопротеида с невыясненной до конца функцией, молекулярная масса которого составляет 50 – 60 кДа (291 аминокислотный остаток; остальная часть молекулы, примерно 1/3 часть по массе, представлена углеводными фрагментами). В настоящее время определена аминокислотная последовательность как a- [9], так b-субъединиц [10], а также установлено расположение полипептидных цепей этих белков в мембране (рис. 2, А). Полипептидная цепь a-субъединицы несколько раз пересекает мембрану, образуя 5 трансмембранных петель. N- и С-концы a-субъединицы находятся в цитоплазме. Большая часть полипептидной цепи (около 800 аминокислот) образует большой цитоплазматический домен, в котором расположен активный центр фермента, где и происходит гидролиз АТФ. Катионы перемещаются через мембрану по каналу, который формируется трансмембранными петлями. N-конец b-субъединицы находится внутри цитоплазмы, ее полипептидная цепь пересекает мембрану только один раз. Большая часть р-субъединицы располагается с внеклеточной стороны мембраны. На ней расположены участки, подвергающиеся гликозилированию.
 и схема, иллюстрирующая каталитический цикл Н<sup>+</sup>, К<sup>+</sup>-АТФазы (Б).")
Рис. 2. Схема укладки a- и b-субъединиц Н+, К+-АТФазы в липидном бислое (А) и схема, иллюстрирующая каталитический цикл Н+, К+-АТФазы (Б).
АТФазы Р-типа осуществляют гидролиз АТФ до АДФ и неорганического фосфата. Высвобождающаяся в процессе гидролиза энергия используется для переноса катионов через мембрану против электро- химического градиента. Характерной особенностью АТФаз Р-типа является образование в процессе каталитического цикла фосфорилированного интермедиата (фосфорилированию подвергается остаток аспарагиновой кислоты, расположенный на a-субьединице; в Н+, Н+, К+-АТФазе это Asp-385). Вторая особенность этого семейства АТФаз заключается в том, что в процессе гидролиза АТФ фермент пребывает в двух основных конформациях – Е1 и Е2, которые различаются по сродству к переносимым катионам. Конформация Е1 имеет высокое сродство к Н+, а конформация Е2 – к катионам К+. Схема гидролиза АТФ Н+, К+-АТФазой представлена на рис. 2, Б. В конформации Е1 со специфическими центрами, расположенными на цитоплазматической поверхности мембраны, связывается H+, после чего происходит фосфорилирование Asp-385, расположенного в активном центре фермента (образование Е1-Р). После фосфорилирования закрываются створки канала, находящиеся на цитоплазматической стороне мембраны. Затем протоны перемещаются через мембрану, что приводит к изменению конформации фермента (переход Е1-Р в Е2-Р). В этом состоянии открываются створки канала с люминальной (внеклеточной) стороны. После этого протоны высвобождаются из катионсвязывающих участков фермента, а К+ связываются с катионсвязывающими центрами на люминальной поверхности мембраны. Связывание К+ с Е2-Р-формой фермента активирует гидролиз ацилфосфатной связи и высвобождение неорганического фосфата. Вслед за этим закрываются створки канала с внеклеточной стороны и ионы калия с внеклеточной поверхности мембраны перемещаются на цитоплазматическую. Связывание АТФ приводит к тому, что происходит изменение конформации фермента (из Е2 переходит в Е1) и К+ высвобождаются в цитоплазму, после чего цикл может повториться. Обмен Н+ на К+, осуществляемый Н+, К+-АТФазой, является электронейтральным. Возникающая в результате работы протонного насоса разница в концентрации Н+ по разные стороны апикальной мембраны составляет 106. Это самый большой градиент концентраций, создаваемый известными в настоящее время системами активного транспорта.
Активность Н+, К+-АТФазы специфически подавляется омепразолом и другими соединениями (лансопразол, пантопразол), являющимися замешенными производными бензимидазола. Эти соединения, накапливаясь в кислых компартментах, главным образом во внутриклеточных канальцах париетальных клеток, связывают Н+ и превращаются в собственно ингибитор, который ковалентно (необратимо) взаимодействует с SH-группами белка, расположенными на люминальной поверхности апикальной мембраны [8]. Восстановление активности Н+, К+-АТФазы после обработки омепразолом происходит главным образом по мере синтеза новых молекул фермента, поэтому длительность вызванного им ингибирования зависит от скорости обновления фермента (половина молекул Н+, К+-АТФазы человека обновляется за 30-48 ч). Кроме того, известны нековалентные (обратимые) ингибиторы Н+, К+-АТФазы [6]. Среди них наиболее изучен имидазопиридин SCH- 28080. Это соединение взаимодействует с К-связывающим участком фермента. Длительность действия этих соединений на Н+, К+-АТФазу зависит главным образом от продолжительности жизни самого соединения, а не фермента.
Кроме Н+, К+-АТФазы, в секреции соляной кислоты участвуют компоненты, обеспечивающие транспорт К+ по градиенту концентраций и сопряженный с ним выход Cl- против градиента концентраций. Транспорт Cl- осуществляется через специальный хлорный канал. В настоящее время этот канал идентифицирован [5]. Он представляет собой белок с молекулярной массой около 100 кДа (898 аминокислот) и по структуре похож на каналы семейства СlС-2, которые присутствуют в мозге и сердце (гомология между хлорным каналом из слизистой оболочки желудка кролика и СlС-2 хлорным каналом из мозга крысы составляет 93%). Проводимость канала равна 7pS при концентрации CsC1 150 мМ с обеих сторон мембраны. Через канал, кроме Cl-, могут проходить и другие анионы. Селективность канала для анионов уменьшается в ряду I-, Cl-, Br-, NO3. Канал является потенциал- и рН-зависимым. Изменение потенциала от 0 до – 80 мВ приводит к 10-кратному увеличению проводимости канала. При потенциале – 80 мВ и внутриклеточном рН 7,4 снижение рН вне клетки до 3,0 дополнительно увеличивает проводимость канала в 5-6 раз.
К+ покидают клетку, по-видимому, через специальный калиевый канал. Установлено, что секрецию соляной кислоты тормозит тетраэтиламмоний, который известен как ингибитор калиевых каналов. Однако в отличие от Cl-канала, структура которого хорошо изучена, калиевый канал идентифицирован только в электрофизиологических экспериментах.
Для выяснения молекулярных механизмов, обеспечивающих активацию секреции соляной кислоты, необходимо выяснить последовательность процессов, происходящих после связывания молекулы секретогена с рецептором, расположенным на поверхности париетальной клетки. В течение многих лет было известно, что париетальная клетка содержит как минимум рецепторы двух типов: гистаминовые H2-рецепторы и мускариновые M3,-рецепторы для ацетилхолина. До недавнего времени не было данных о рецепторе для гастрина. Считалось, что гастриновые рецепторы находятся на энтерохромаффинных клетках, которые после связывания гастрина высвобождают гистамин. Выделяющийся из энтерохромаффинных клеток гистамин связывается с H2-рецепторами париетальных клеток, обеспечивая стимуляцию секреции. Однако недавно было показано, что и париетальные клетки содержат гастриновые рецепторы [11]. Рецептор для гастрина относится к типу В-рецепторов для холецистокинина (ССК-В). Рецепторы этого типа, как и находящиеся на поверхности париетальных клеток М,-рецепторы, обеспечивают свое действие через О-белки, активирующие фосфолипазу С. Этот фермент гидролизует фосфоинозитиды, находящиеся в липидном слое мембраны. По-видимому, образующийся в результате гидролиза инозитолтрифосфат вызывает выход Са2+ из внутриклеточных депо (эндоплазматический ретикулум), в результате чего внутриклеточная концентрация Са2+ увеличивается. Второй продукт этой реакции диацилглицерол вместе с Са2+ активирует Са, фосфолипидозависимую протеинкиназу (протеинкиназа С), которая в свою очередь фосфорилирует белки мишени, влияя на их функциональную активность. Таким образом, в результате активации париетальных клеток под действием как гастрина, так и ацетилхолина могут происходить увеличение внутриклеточной концентрации Са2+ и фосфорилирование белков-мишеней под действием протеинкиназы С. Однако вся цепь событий для париетальной клетки не прослежена. Известно лишь, что активация секреции соляной кислоты париетальными клетками под действием гастрина и ацетилхолина приводит к повышению концентрации вторичного мессенджера цГМФ. Возможные начальные этапы процесса активации секреции париетальными клетками представлены на рис. 3.
Наиболее изучена активация секреции соляной кислоты под действием гистамина. Связываясь с H2- рецептором, этот секретоген через О-белки активирует аденилатциклазу, в результате чего повышается внутриклеточный уровень цАМФ [1]. Вслед за этим происходит повышение внутриклеточной концентрации Са2+: он входит в клетку через плазматическую мембрану. Париетальные клетки содержат цАМФ-зависимые протеинкиназы (протеинкиназы А) двух типов – I и II. Установлено, что мишенями для цАМФ-зависимых протеинкиназ является большое количество как цитоплазматических, так и мембранных белков. Одной из идентифицированных мишеней протеинкиназы А является Cl-канал. В системе in vitro установлено, что фосфолирование канала этой протеинкиназой приводит к увеличению его проводимости. В экспериментах с мембранными везикулами, полученными из стимулированных гистамином и несекретирующих (обработанных антагонистом H2-рецептора циметидином) париетальных клеток, было показано, что скорость секреции соляной кислоты в несекретирующих клетках лимитируется не активностью Н+, К+-АТФазы, а проницаемостью Cl-канала. Таким образом, фосфорилирование Cl-канала протеинкиназой А устраняет лимитирующую стадию в процессе секреции соляной кислоты.
 и возможные механизмы активации")
Рис. 3. Рецепторы, через которые осуществляется активация секреции в париетальной клетке (схема) и возможные механизмы активации
В несекретирующих париетальных клетках большая часть Н+, К+-АТФазы является неактивной и сосредоточена в везикулах, расположенных в цитоплазме неподалеку от апикальной поверхности мембраны (так называемые тубуловезикулы). Активация секреции сопряжена в первую очередь с перемещением этих везикул к поверхности апикальной мембраны или мембраны канальцев и с их слиянием с этими мембранами. Этот процесс сопровождается увеличением количества молекул Н+, К+-АТФазы на единицу поверхности мембраны. Активное участие в этом процессе принимает цитоскелет париетальной клетки: обработка клеток цитохалазинами А и Е, которые блокируют удлинение микрофиламентов, предотвращает активацию секреции. Известно, что около 4% белка париетальной клетки представлено актином и около 60% актина находится в полимеризованной F-форме [3]. Нити полимеризованного актина, сшитые друг с другом специальными белками цитоскелета, располагаются внутри микроворсинок апикальной поверхности мембраны, формируя своеобразный скелет. Другие белки цитоскелета сшивают нити актина с белками, встроенными в мембрану. Активация секреции сопряжена с перемещением актина к поверхности апикальной мембраны, а миозина – ближе к центру клетки. По-видимому, среди фосфорилируемых протеинкиназой А белков париетальных клеток немало белков цитоскелета. В частности, среди мишеней протеинкиназы А идентифицирован периферический белок мембраны эзрин с молекулярной массой 80 кДа, который участвует в связывании актиновых микрофиламентов с мембраной [4].
При стимуляции клетки гистамином наблюдается перераспределение белков между цитоплазмой и мембраной. Удалось обнаружить белки, которые при активации секреции перемещаются из цитозоля в мембрану и специфически взаимодействуют с Н+, К+-АТФазой. Известно, что Н+, К+-АТФаза, как и другие АТФазы P-типа, ингибируется мелитгиномпептидом из яда пчелы, состоящим из 26 аминокислот. Имеющиеся в настоящее время данные позволяют предполагать, что мелиттин имитирует определенную детерминанту, участвующую в белок-белковых взаимодействиях в клетке. При использовании антител на мелиттин в цитозоле несекретирующих париетальных клеток был обнаружен белок с молекулярной массой 67 кДа. Этот белок взаимодействует с антителами на мелиттин и, следовательно, содержит участки, по структуре похожие на мелиттин. При стимуляции париетальных клеток гистамином мелиттиноподобный белок перемещается из цитозоля в мембрану [2]. Белок был получен в чистом виде, и в экспериментах in vitro установлено, что он специфически взаимодействует с Н+, К+-АТФазой. Однако пока неизвестно, что является сигналом для перемещения мелиттиноподобных белков при стимуляции, а также какова их функция. Не исключено, что они также являются белками цитоскелета, осуществляющими взаимодействие Н+, К+-АТФазы с микрофиламентами. Полученная в последние годы информация позволяет считать, что белки, обеспечивающие связывание мембранных белков с цитоскелетом, зачастую не только являются структурным элементом, но и участвуют в передаче сигнала. Таким образом, мелиттиноподобный белок, перемещающийся в мембрану при стимуляции секреции, может оказаться и активатором Н+, К+-АТФазы.
Завершая обзор экспериментальных данных, можно сделать следующее заключение: несмотря на то что представление о молекулярных механизмах активации секреции соляной кислоты значительно расширилось, детальное изучение этого процесса лишь начинается и в ближайшее время можно ожидать появления новых интересных фактов.
Список литературы
1. Cuppoletti У., Malinowska D., Sachs G. Biochim. biophys. Acta. – 1988. – Vol. 972. – P.95 – 105.
2. Cuppoletti Х., Huang Н., Kaetzel M, Malinowska D. Amer.J.Physiol. – 1993. – Vol. 264.- P. 637 – 644.
3. Dabihe M, Munizaga А., Koenig С.S. Biol.Res. – 1994. – Vol. 27. – P.29 – 38.
4. Hanzel D., Reggio Н., Bretcher А. et al. ЕМВО Journ.- 1991. – Vol. 10. – P.2363-2373.
5. Malinowska D.Н, Kupert ТХ, Baninski А. et al. Amer. J. Physiol. – 1995. – Vol. 268. – P. 191 – 200.
6. Мope А., Sachs G. Biochem. Soc. Trans. – 1992. – Vol. 20.- P. 566-572.
7. Radon Е.С., Reuben М.А. Ann. Rev.Physiol. – 1990.-Vol. 52. – P. 321 – 344.
8. Sachs G., Shin J;М, Briving С. et al. Ann. Rev. Pharmacol. Toxicol. – 1995. – Чо1. 35. – P. 277 – 305.
9. Shull G.Е., Етое! I.В. J. biol. Chem. – 1986. – Vol. 261. – P. 16788-16791.
10. Shull G.Е. J.biol. Chem. – 1990. – Vol. 265. – P.12123-12126.
11. Wolfe M.M, 7seny С.С. Gastroenterology. – 1993.-Vol. 104. – P. 1876 – 1878.
Молекулярные механизмы регуляции секреции соляной кислоты слизистой оболочки желудка.
О.Д. Лопина, А.А. Котлобай, А.М. Рубцов.
(Кафедра биохимии биологического факультета Московского государственного университета им. М.В. Ломоносова).
Российский журнал гастроэнтерологии, гепатологии, колопроктологии. 1997, №6, с. 15-19.
Источник