Гладкие мышцы желудка сокращаются

Гладкие мышцы желудочно-кишечного тракта. Виды сокращения мышц желудочно-кишечного тракта
а) Электрическая активность гастроинтестинальных гладких мышц. В результате собственной медленной электрической активности, распространяющейся вдоль мембран мышечных волокон, происходит почти непрерывное возбуждение гладкой мускулатуры желудочно-кишечного тракта. Эта активность имеет два основных типа электрических волн:
(1) медленные волны;
(2) распространяющийся потенциал.
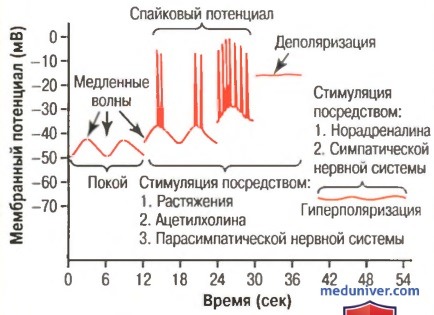
Мембранный потенциал гладкой кишечной мускулатуры кишечника. Отмечены медленные волны, спайковые потенциалы, общая деполяризация и гиперполяризация. Все это происходит в результате различных физиологических состояний кишечника
Величина мембранного потенциала покоя гладкой мышцы пищеварительной системы может колебаться, и это имеет важное значение в управлении моторной активностью желудочно-кишечного тракта.
б) Медленные волны. В большинстве случаев сокращение желудка и кишечника происходят ритмично, и этот ритм устанавливается в основном частотой так называемых медленных волн мембранного потенциала гладкой мускулатуры. Эти волны, показанные на рис. 62-3, не являются потенциалами действия. Они представляют собой медленные волнообразные изменения мембранного потенциала покоя.
Их интенсивность обычно колеблется между 5 и 15 мВ, а частота их возникновения в различных отделах желудочно-кишечного тракта человека варьирует от 3 до 12 в минуту: около 3 — в теле желудка, до 12 — в двенадцатиперстной кишке и 8-9 — в конечной части подвздошной кишки. Следовательно, частота сокращений тела желудка составляет обычно около 3 сокращений в минуту, двенадцатиперстной кишки — приблизительно 12 в минуту, подвздошной кишки — 8-9 в минуту.
Конкретные причины появления медленных волн до конца не установлены. Возможно, они возникают в результате действий комплекса взаимных влияний между клетками гладкой мускулатуры и специальных клеток, называемых интерстициальными клетками Кахаля, которые, по-видимому, являются электрическими пейсмекерами для гладкомышечных клеток. Эти интерстициальные клетки формируют сеть и расположены между слоями гладких мышц. Они сообщаются с гладкой мускулатурой посредством синаптических контактов.
В интерстициальных клетках Кахаля благодаря уникальным ионным каналам, которые периодически открываются и генерируют входящий (пейсмекерный) ток, происходят циклические изменения величины мембранного потенциала — т.е. генерация медленных волн.
Медленные волны обычно не вызывают сокращений гладкой мускулатуры в большинстве отделов желудочно-кишечного тракта, за исключением желудка. Вместо этого они вызывают в основном появление ритмических спайков, которые, в свою очередь, вызывают мышечное сокращение.
в) Спайковые потенциалы. Спайковые потенциалы действуют как потенциалы действия. Это происходит автоматически, когда мембранный потенциал покоя гладкомышечной клетки становится более положительным и составляет -40 мВ (обычное значение для мембранного потенциала покоя волокон гладкой мускулатуры желудочно-кишечного тракта колеблется от -50 до -60 мВ).
На рисунке выше показано, что каждый раз, когда пик медленных волн на время достигает отметки -40 мВ и выше, появляются спайковые потенциалы.
Чем выше поднимаются медленные волны, тем с большей частотой возникают спайковые потенциалы; обычно они варьирует между 1 и 10 спайками в секунду. Спайковый потенциал в мышцах желудочно-кишечного тракта длится в 10-40 раз дольше, чем потенциал действия в крупных нервных волокнах. Каждый потенциал действия пищеварительной системы длится от 10 до 20 мсек.
Другое важное отличие между потенциалами действия гладкой мускулатуры кишечника и потенциалами действия нервных волокон заключается в механизме их возникновения. В нервном волокне потенциал действия возникает в результате стремительного проникновения ионов натрия через натриевые каналы во внутреннюю среду волокна.
В гладкой мускулатуре кишечника каналы, отвечающие за потенциал действия, несколько иные: они позволяют проходить в основном большому количеству ионов кальция и небольшому количеству ионов натрия. Их называют кальций-натриевыми каналами. Эти каналы открываются и закрываются гораздо медленнее, чем быстрые натриевые каналы нервных волокон. Медленная скорость закрытия и открытия кальций-натриевых каналов является причиной большой продолжительности потенциала действия. Движение большого количества ионов кальция во внутреннюю среду мышечного волокна во время потенциала действия также играет особую роль в сокращении мышечных волокон, о чем было сказано ранее.
– Также рекомендуем “Потенциал покоя мышц желудочно-кишечного тракта. Тоническое сокращение мышц кишечника”
– Вернуться в оглавление раздела “Физиология человека.”
Оглавление темы “Физиология мозгового кровотока”:
1. Регуляция мозгового кровотока. Саморегуляция мозгового кровотока
2. Микроциркуляция мозга. Причины инсультов
3. Система цереброспинальной жидкости. Функции спинномозговой жидкости
4. Секреция спинномозговой жидкости. Физиология периваскулярных пространств
5. Давление спинномозговой жидкости. Измерение давления спинномозговой жидкости
6. Причины отека соска зрительного нерва. Причины гидроцефалии
7. Гематоэнцефалический барьер. Отек мозга
8. Метаболизм мозга. Регуляция метаболизма мозга
9. Физиология желудочно-кишечного тракта. Двигательная активность желудочно-кишечного тракта
10. Гладкие мышцы желудочно-кишечного тракта. Виды сокращения мышц желудочно-кишечного тракта
Источник
Оглавление темы “Глотание. Пищеварение в желудке.”: Сократительная деятельность мускулатуры желудка. Сокращение желудка. Работа желудка.Во время приема пищи происходит расслабление мускулатуры фундального отдела желудка, что является рефлекторной реакцией в ответ на раздражение рецепторов слизистой оболочки полости рта, глотки и пищевода. Это расслабление получило название рецептивной релаксации, которая способствует заполнению пищей фундального отдела. Через 5—30 мин после завершения приема пищи происходит усиление моторики желудка. Вначале автоматическое возбуждение миоцитов возникает в водителе ритма, который располагается в области кардии, откуда волна перистальтического сокращения распространяется в дистальном направлении — на тело желудка, его антральную часть и пилорический сфинктер. За 1 мин возникает около трех волн.
Скорость распространения перистальтической волны по мере ее приближения к пилорическому отделу желудка возрастает с 1 до 3—4 см/с. Волна перистальтики представляет собой сокращение участков (полос) циркулярного слоя мышц по всей окружности желудка. После сокращения участка циркулярных мышц происходит их расслабление, а волна сокращения переходит на другой участок. Перистальтические волны смещают в дистальном направлении слои химуса, прилежащие к слизистой оболочке желудка, в наибольшей степени обработанные желудочным соком. На фоне перистальтических волн возникают сильные и продолжительные сокращения (тонические волны), которые смещают химус из фундального отдела в пилорический. По мере накопления содержимого желудка в пилорическом отделе возникают сильные систолические сокращения его мускулатуры. Они создают значительное возрастание давления химуса в желудке, что при открытом пилорическом сфинктере вызывает эвакуацию порции желудочного содержимого в двенадцатиперстную кишку.
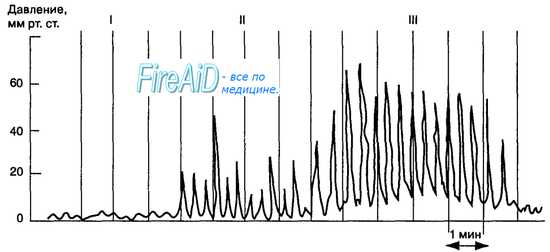
Если с помощью введенного в желудок человека баллона регистрировать колебания давления желудочного содержимого, то можно выявлять три типа волн сокращения (рис. 11.13): фазные волны с амплитудой до 5 мм рт. ст., продолжительностью около 20 с, частотой 3 в 1 мин (I тип), фазные волны той же продолжительности и частоты, но большей амплитуды (10—50 мм рт. ст. — II тип). На фоне волн I и II типов происходит возрастание исходного тонуса мускулатуры, что повышает давление на 5— 10 мм рт. ст. в течение 5—6 мин (III тип). – Вернуться в оглавление раздела “Физиология человека.” |
Источник
Оглавление темы “Функции пищеварительной системы ( жкт ). Типы пищеварения. Гормоны желудочно-кишечного тракта. Моторная функция желудочно-кишечного тракта.”:
1. Физиология пищеварения. Физиология пищеварительной системы. Функции пищеварительной системы ( жкт ).
2. Состояние голода и насыщения. Чувство голода. Чувство насыщения. Гиперфагия. Афагия.
3. Секреторная функция пищеварительной системы. Секреция. Секрет пищеварительных желез.
4. Типы пищеварения. Собственный тип пищеварения. Аутолитический тип. Внутриклеточное пищеварение. Внеклеточное пищеварение.
5. Гормоны желудочно-кишечного тракта. Место образования гормонов жкт. Эффекты вызываемые гормонами желудочно – кишечного тракта.
6. Моторная функция желудочно-кишечного тракта. Гладкая мускулатура пищеварительного тракта. Cфинктеры жкт. Сократительная деятельность кишечника.
7. Координация сократительной деятельности. Медленные ритмические колебания. Продольный мышечный слой. Влияние катехоламинов на миоциты.
Моторная функция желудочно-кишечного тракта. Гладкая мускулатура пищеварительного тракта. Cфинктеры жкт. Сократительная деятельность кишечника.
Процесс пищеварения во всех отделах пищеварительного тракта осуществляется при участии двигательной активности его мускулатуры. Сокращения мышц обеспечивают: прием и измельчение пищи в процессе жевания в ротовой полости, глотание и продвижение порции пищи по пищеводу, накопление ее в желудке и эвакуацию его содержимого в кишечник, сокращение и расслабление желчного пузыря, перемешивание и продвижение кишечного содержимого, движение ворсинок, переход химуса из тонкой кишки в толстую, его перемещение по толстой кишке, сокращение и расслабление сфинктеров, перистальтику выводных протоков пищеварительных желез и выведение экскрементов.
Гладкая мускулатура пищеварительного тракта состоит из гладкомы-шечных клеток (миоцитов). Они собраны в пучки и соединены друг с другом нексусами. Пучок получает нервные терминали, артериолу и выполняет роль функциональной единицы гладкой мышцы. Миоциты обладают способностью к спонтанному ритмическому возбуждению за счет периодической деполяризации их мембраны. Это возбуждение распространяется благодаря нексусам от клетки к клетке (как по синцитию). Пучки миоцитов образуют гладкомышечные слои пищеварительной трубки — циркулярный (внутренний), продольный (наружный) и подслизистый (косой).
Растяжение мышц содержимым желудочно-кишечного тракта является для них адекватным раздражителем, вызывающим деполяризацию мембран их клеток и сокращение мышечных волокон. Частота и сила сокращений миоцитов изменяются в широком диапазоне под влиянием нервных импульсов эфферентных терминалей вегетативных нервных волокон, гормонов и гастроинтестинальных регуляторных пептидов. Комплексная нервно-гуморальная регуляция миоцитов обеспечивает соответствие уровня активности мускулатуры объему и составу содержимого желудка и кишечника.
Характер сократительной деятельности мускулатуры пищеварительного тракта зависит от активности водителей ритма, расположенных в желудке и кишечнике. Они представляют собой гладкомышечные клетки, более чувствительные к биологически активным веществам и имеющие более обильную иннервацию, чем другие пучки миоцитов.
На протяжении пищеварительного тракта у человека имеется около 35 сфинктеров. Они состоят из мышечных пучков, расположенных циркулярно (в основном), спирально и продольно. Сокращение циркулярных пучков приводит к смыканию сфинктера, а сокращение спиральных и продольных пучков увеличивает его просвет, что способствует переходу содержимого пищеварительного тракта в нижележащий отдел. Сфинктеры обеспечивают движение содержимого пищеварительной трубки в каудальном направлении и временное разобщение функционально различных частей пищеварительного тракта. Основные из них — кардиальный (на входе в желудок), пилорический (на выходе из желудка), в основании баугиниевои заслонки (на входе в слепую кишку), внутренний и наружный анальный (на выходе из прямой кишки).
– Также рекомендуем “Координация сократительной деятельности. Медленные ритмические колебания. Продольный мышечный слой. Влияние катехоламинов на миоциты.”
Источник
Механизм сокращения гладкой мышцы. Химические основы сокращений гладкой мышцыГладкие мышцы содержат актиновые и миозиновые нити, имеющие химические характеристики, подобные актиновым и миозиновым нитям скелетных мышц. Но в гладких мышцах нет тропонинового комплекса, необходимого для запуска сокращения скелетной мышцы, следовательно, механизм инициации сокращения в них другой. Этот механизм подробно обсуждается далее в нашей статье. Химические исследования показали, что актиновые и миозиновые нити, извлеченные из гладких мышц, взаимодействуют друг с другом во многом так же, как и в скелетной мышце. Более того, процесс сокращения активируется ионами кальция, а энергия для сокращения обеспечивается разрушением АТФ до АДФ. Существуют, однако, значительные различия в морфологической организации гладких и скелетных мышц, а также в сопряжении возбуждения и сокращения, механизме запуска ионами кальция сократительного процесса, длительности сокращения и количестве энергии, необходимой для сокращения.
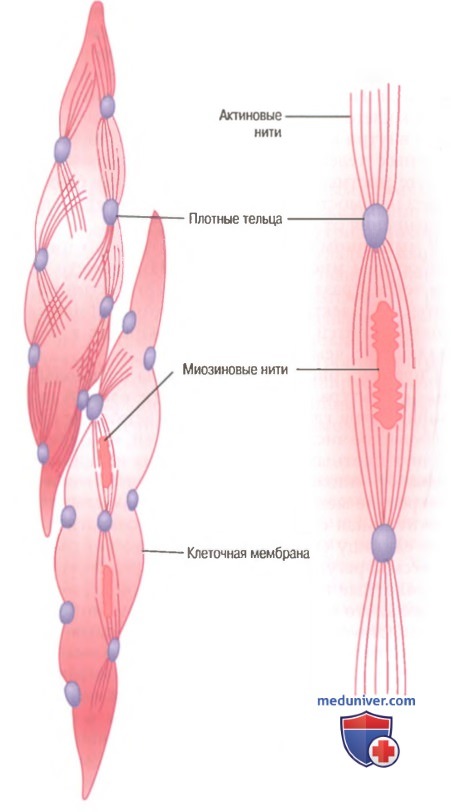
Морфологическая основа сокращения гладких мышцГладкие мышцы не имеют такой упорядоченной организации актиновых и миозиновых нитей, которая обнаруживается в скелетных мышцах, придавая им «полосатость». С помощью техники электронной микрофотографии выявляется гистологическая организация. Видно большое число актиновых нитей, прикрепленных к так называемым плотным тельцам. Некоторые из этих телец прикрепляются к клеточной мембране, другие распределяются внутри клетки. Некоторые из мембранных плотных телец соседних клеток связываются вместе мостиками из внутриклеточных белков. Через эти мостики в основном передается сила сокращения от одной клетки к другой. В мышечном волокне среди актиновых нитей разбросаны миозиновые нити. Их диаметр более чем в 2 раза превышает диаметр актиновых нитей. На электронных микрофотографиях актиновых нитей обычно обнаруживают в 5-10 раз больше, чем миозиновых.
На рисунке представлена предполагаемая структура отдельной сократительной единицы внутри гладкомышечной клетки, где видно большое число актиновых нитей, исходящих от двух плотных телец; концы этих нитей перекрывают миозиновую нить, расположенную посередине между плотными тельцами. Эта сократительная единица похожа на сократительную единицу скелетной мышцы, но без специфической регулярности ее структуры. В сущности, плотные тельца гладкой мышцы играют ту же роль, что и Z-диски в скелетной мышце. Существует и другое различие. Большинство миозиновых нитей имеют поперечные мостики с так называемой боковой полярностью. Мостики организованы следующим образом: на одной стороне они шарнирно фиксируются в одном направлении, а на другой — в противоположном направлении. Это позволяет миозину тянуть актиновую нить с одной стороны в одном направлении, одновременно продвигая с другой стороны другую актиновую нить в противоположном направлении. Такая организация позволяет гладкомышечным клеткам сокращаться с укорочением до 80% их длины вместо укорочения менее чем на 30%, характерного для скелетной мышцы. Большинство скелетных мышц сокращаются и расслабляются быстро, но сокращения гладких мышц в основном являются длительными тоническими сокращениями, которые иногда продолжаются в течение нескольких часов или даже дней. Следовательно, можно ожидать, что морфологические и химические особенности гладких мышц должны отличаться от соответствующих характеристик скелетных мышц. Далее обсуждаются некоторые из этих отличий. Медленная циклическая активность миозиновых поперечных мостиков. В гладкой мышце по сравнению соскелетной гораздо меньше скорость циклической активности миозиновых поперечных мостиков, т.е. скорость их прикрепления к актину, отсоединение от актина и повторное прикрепление для осуществления следующего цикла. Фактически частота циклов составляет лишь от 1/10 до 1/300 этого показателя в скелетной мышце. Однако, как считают, в гладкой мышце значительно больше относительное количество времени, в течение которого поперечные мостики остаются прикрепленными к актиновым нитям, что является главным фактором, определяющим силу сокращения. Возможной причиной медленного циклирования является гораздо меньшая по сравнению со скелетной мышцей АТФ-азная активность головок поперечных мостиков, в связи с чем скорость разрушения АТФ — источника энергии для движения головок поперечных мостиков — значительно снижена с соответствующим замедлением скорости их циклов. – Также рекомендуем “Энергообеспечение сокращения гладкой мышцы. Механизм защелки гладкой мышцы” Оглавление темы “Механизмы сокращения мышечных клеток. Сокращение гладких мышц”: |
Источник